Improved development of mouse and human embryos using a tilting embryo culture systemRBM Online28 March 2010 |
Koji Matsuura a,b, Nobuyoshi Hayashi c, Yuka Kuroda a,b, Chisato Takiue c, Rei Hirata c, Mami Takenami a, Yoko Aoi c, Nanako Yoshioka c, Toshihiro Habara c, Tetsunori Mukaida d, Keiji Naruse a,*
a Cardiovascular Physiology, Graduate School of Medicine, Dentistry and Pharmaceutical Sciences, Okayama University, Okayama, Japan; b Research Core for Int erdisciplinary Sciences, Okayama University, Okayama, Japan; c Okayama Couples Clinic, Okayama, Japan; d Hiroshima HART Clinic, Hiroshima, Japan
* Corresponding author.
E-mail address: [email protected] (K Naruse).

Dr Keiji Naruse graduated from Nagoya University School of Medicine in 1988 (MD) and received his PhD in medicine from Nagoya University in 1992. He was an associate professor at Nagoya University from 1999 to 2005 and is currently a chairman and professor of Cardiovascular Physiology at Okayama University Graduate School of Medicine. He was a visiting professor at Harvard Medical School from 1998 to 2001. He has been working in the fields of mechanobiology of circulation, reproduction, and sensory systems.
Abstract
Mammalian embryos experience not only hormonal but also mechanical stimuli, such as shear stress, compression and friction force in the Fallopian tube before nidation. In order to apply mechanical stimuli to embryos in a conventional IVF culture system, the tilting embryo culture system (TECS) was developed. The observed embryo images from the TECS suggest that the velocities and shear stresses of TECS embryos are similar to those experienced in the oviduct. Use of TECS enhanced the development rate to the blas- tocyst stage and significantly increased the cell number of mouse blastocysts (P < 0.05). Although not statistically significant, human thawed embryos showed slight improvement in development to the blastocyst stage following culture in TECS compared with static controls. Rates of blastocyst formation following culture in TECS were significantly improved in low-quality embryos and those embryos cultured under suboptimal conditions (P < 0.05). The TECS is proposed as a promising approach to improve embryo development and blastocyst formation by exposing embryos to mechanical stimuli similar to those in the Fallopian tube.
RBMOnline © 2009, Reproductive Healthcare Ltd. Published by Elsevier Ltd. All rights reserved.
Declaration: Keiji Narusee launched the bio-venture company, Strex Inc., in 2003 and serves as a Chief Scientific Officer. The other authors report no financial or commercial conflicts of interest.
KEYWORDS: blastocyst, embryo development, mechanical stimuli, shear stress, tilting embryo culture system
Introduction
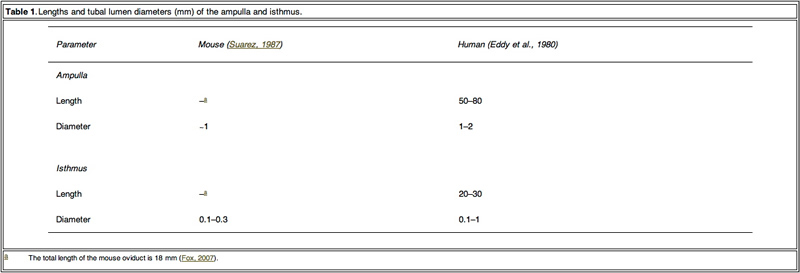
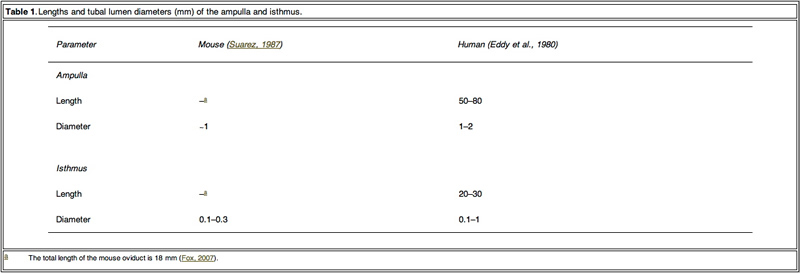
Mammalian embryos are transported to the uterine cavity through the Fallopian tube during cell cleavage, blastomere and blastocyst development (Eddy and Pauerstein, 1980; Halbert et al., 1976). In conjunction with ciliated epithelium, the Fallopian tube acts as a peristaltic pump due to phasic contraction of the smooth muscle in the wall to transport the embryo (Lyons et al., 2006; Zervomanolakis et al., 2007). Based on the movement of cilia and the similar size between the tubal lumen of ampulla and isthmus (Table 1) and the diameter of the embryo (0.1 mm), fertilized oocytes may be subject to a mechanical influence from the Fallopian tube lumen, such as compression and shear stress from the tubal fluid. It is postulated that these mechanical actions in the Fallopian tube might play an important role in embryo development. However, conventional in-vitro static culture conditions do not mimic mechanical stimuli to embryos. Providing mechanical stimuli to developing embryos in-vitro, similar to that experienced in the Fallopian tube, may improve development.
From the 1980s, some groups reported in-vitro non-static culture results (Nagai et al., 1993; Staigmiller and Moor, 1984). However, these reports did not discuss the precise effect on embryo development from cleaved embryo to blastocyst. Recent papers have reported that a microfluidic dynamic embryo culture system with media flow improved mouse embryo development (Cabrera et al., 2006). The report proposed the importance of the physical/mechanical environment on embryo development. Because the systems are complex to handle, clinical application of the culture system is quite difficult. In addition to ease-of-use, another consideration regarding mechanical stimulation during embryo development is avoiding excess stress. Excess mechanical stimuli would damage embryos. It was found that shear stress over 1.2 dyn/cm2 caused lethality within 12 h for blastocysts (Xie et al., 2006). Over-handling of embryos, such as excess pipetting, caused elevation of phosphorylated stress- activated protein kinase and may cause rapid transient changes in hundreds of proteins and mRNA (Xie et al., 2007).
In order to construct a dynamic culture system that can apply a mechanical stimulus and can be easily adapted to conventional static culture platforms, the tilting embryo culture system (TECS) was developed. Placing a conventional culture dish or plate on a tilted plate makes embryos in culture move along the bottom of the dish by gravity. Animal model experiments using TECS were first conducted, for the in-vitro culture of mouse 2-cell-stage embryos. A subsequent study using thawed human embryos cultured on TECS was also performed.
Materials and methods
Observation of embryo motion and shear stress calculation
To estimate the shear stress applied to embryos, mouse embryo motion in the microdrop on the tilting plate of a prototype TECS was observed by objective lens (20x) attached to the bottom of the tilting TECS plate. To estimate the flow velocity of media in the microdrop, this study observed the motion of the microspheres (0.5–5 μm diameter) in the microdrop covered with mineral oil by tilting inverted microscope (Eclipse, Nicon, Tokyo, Japan), particularly focusing on the motion of microspheres at the centre of the bottom where embryos in the microdrop were moving. When the microscope was tilted, the particles moved in the same direction as the leaning microscope. The average particle velocities, which were considered as the velocity of the medium in the microdrop, were calculated. Under the holding the tilt and static conditions, Brownian motion of the particles was dominant. Images of moving embryos and microspheres were recorded by a charge-coupled device camera connected to a personal computer. The frame rate of the recording was 30 frames/s.
The observed maximum velocity of the embryos and the velocity of the medium were VE and VM, respectively. The velocity to estimate maximum shear stress (VSS) was calculated from the difference between VE and VM.
VSS =VE -VM
The maximum shear stress applied to embryos during the tilting was calculated from the following equation:
Shear stress = 6µrVSS =4µr2 = 6VSS dyn/cm2
where µ is the viscosity of the medium and r is the radius of the embryo.
In an attempt to provide as accurate an estimate as possible regarding sheer stress experienced by embryos, this study attempted to determine the flow velocity of fluids using microspheres. The above equations are applicable in the flow condition. Generally, the effect of Brownian motion during flow can be neglected for the shear stress calculation. However, it would be difficult to determine accurate fluid velocity by particle image velocimetry. Although forces such as friction come into play with this approach, it provides a rough estimation for the final calculations.
Embryo motion in extirpated mouse oviducts was observed as follows. Female ICR mice (8–12weeks old; Charles River Japan, Yokohama, Japan and Shimizu Laboratory Supplies, Kyoto, Japan) were injected with 5 IU pregnant mare’s serum gonadotrophin (Aska Pharmaceutical, Tokyo, Japan), followed by 5 IU human chorionic gonadotrophin (Aska Pharmaceutical) 48h later and mated with males. Successful mating was determined the following morning by the detection of a vaginal plug. Two days after the mating, the oviduct was extirpated with embryos and the oviduct was sandwiched between a cover slip and a glass slide to observe the embryo in the oviduct. The velocity of the embryo in extirpated mouse oviducts was calculated from the movie of the embryo motion. Animal use protocols were approved by Okayama University animal investigation committee.
Specification and motion programmes of TECS
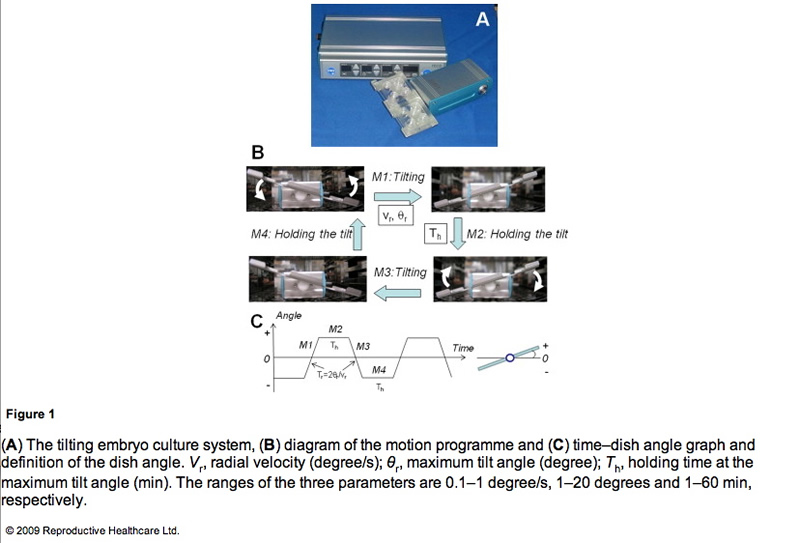
TECS (Strex, Osaka, Japan) is an electrical device with a power cord that is designed to be used in a humidified incubator. This device consists of a control unit (Figure 1A, upper) and a motor unit with a tilting plate (Figure 1A, lower). Four-well chambers and/or dishes are set on the plate. The TECS motor unit is controlled and DC-powered by a cord connected to the control unit through an access hole in the incubator. If the incubator does not have an access hole, a flat-type cable extends tightly through the door of the incubator in order not to alter temperature, gas concentration and humidity inside the incubator. The motor unit is waterproof. The controller outside the incubator puts in the motion parameters.

A representative motion programme of TECS is shown in Figure 1B and C. The TECS can control the parameters of the uniform radial velocity (Vr), the maximum tilt angle (θr) and the holding time at the maximum tilt angle (Th). The tilting time (Tr) in seconds is calculated from 2θr/Vr. First, the plate is tilted to the positive maximum tilt angle (+θr) (M1). Second, the tilting plate is held for Th with no motion (M2). Third, the plate is tilted to the negative maximum tilt angle (−θr) (M3). Last, the tilting plate is held for Th with no motion (M4). The TECS motion cycle is summarized in Figure 1C. The cycle continues until the power is turned off.
To apply conventional culture dishes/approaches to the TECS, spill out of the mineral oil should be prevented. The angles at which mineral oil begins to spill out are different according to the size of the dishes. The mouse embryo experiments used dishes of 35 mm diameter and maximum tilt of 20 degrees. However, in the human embryo study, the plate could not be tilted over 10 degrees due to the use of dishes of 60 mm diameter.
The maximum tilt angle that caused mineral oil to spill out was surveyed and the limit of the maximum tilt angle was approximately 20 degrees in the microdrop setup. Furthermore, excess uniform radial velocity also induces spill out of the mineral oil. The minimum radial velocity at which the mineral oil spilt from the 35 mm dish was 240 degree/s when the maximum tilt angle was 20 degrees. The oil did not spill out when the tilt angle was 10 degrees. It is necessary to increase the maximum tilt angle and radial velocity in order to move embryos in the microdrop. However, this study set the maximum tilt angle and radial velocity so that they did not result in the spilling out of the mineral oil but still allowed the observation of embryo motion in the video rate recording. Thus, the maximum tilt angle was 10–20 degrees and the radial velocity was approximately 1 degree/s.
Tilting mouse embryo culture
Frozen 2-cell-stage embryos of ICR mouse (Arc Resources, Kumamoto, Japan) were thawed and cultured in 50 and 500 μl, respectively, of potassium simplex optimized embryo culture medium (Daiya Shiyaku, Tokyo, Japan). The culture medium was covered with mineral oil and incubated for 3 days in a humidified environment of 5% CO2 in air at 37°C. In the TECS group, the mouse embryos were cultured at a maximum tilt angle of 20 degrees with a holding time of 1 min. The plate was rotated at 1 degree/s to reach a total tilt of 20 degrees. To count cells in the mouse blastocyst, the cells were stained with Hochest 33342, as previously reported (Hardy et al., 1989; Mottla et al., 1995). The stained blastocysts were observed by confocal microscopy (FV-1000; Olympus, Tokyo, Japan) and a 3D image was constructed.
Human embryo study
The human embryo study used 3- to 11-cell-stage embryos frozen by the slow method 3 days after collection of the oocytes from May 2000 to December 2004 (Cohen et al., 1985) and extended the prospective study of 220 fertilized human embryos that were to be discarded with consent after pregnancy of patients. This study was approved by the ethics committee of the Okayama University Graduate School of Medicine.
The frozen embryos were thawed with THAW-KIT 1 (Vitrolife, Gothenburg, Sweden) and the viability of the thawed embryos was approximately 80%. Once thawed, human embryos were cultured in a 20 ll microdrop of Global medium (LifeGlobal, Ontario, Canada) covered with mineral oil for 48 h. The thawed embryos with morphologically regular cleavage were divided so that there would be the same percentage of embryos with regular cleavage (33%) in both the TECS group and the control group. The viable cell number of the thawed human embryos was from three to 11. The dishes were placed on the tilting plate of TECS in a 50 l multi-gas incubator (ASTEC, Fukuoka, Japan). The thawed human embryos were cultured at a 10 degree tilt for 10 min. The radial velocity of the tilt was 1 degree/s (Vr).
The blastocysts were morphologically evaluated according to the classification of Gardner et al. (2000). The cells in the human blastocyst were stained with Hochest 33342, the blastocysts were observed by confocal microscopy (FV-1000 Olympus, Tokyo, Japan) and a 3D image was constructed.
Statistical analysis
Chi-squared test and Student’s t-test were used to determine differences in the blastocyst development rate and in the number of cells in the blastocysts between the groups, respectively. A P-value < 0.05 was considered significant.
Observation of embryos in motion by TECS and estimation of shear stress
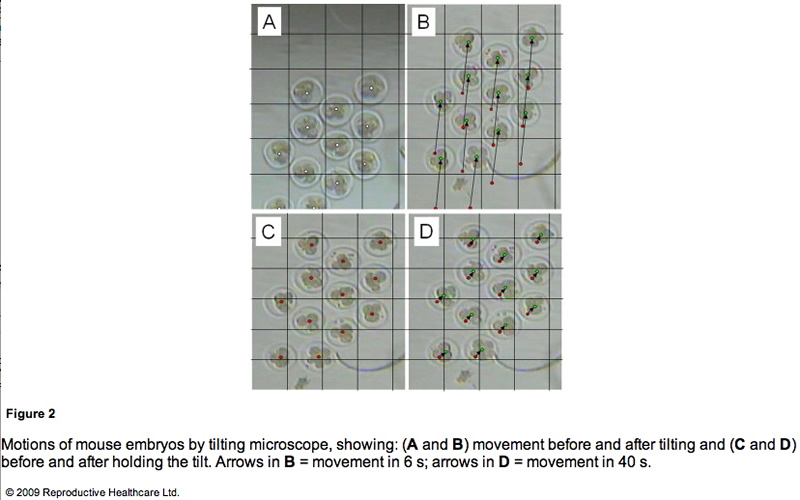
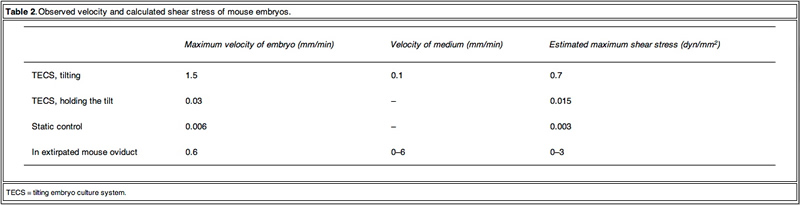
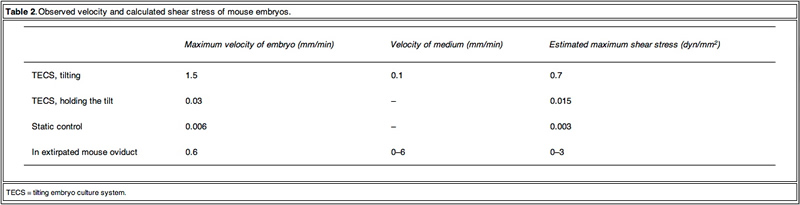
To obtain detailed information on the embryo motion, the blastomeres were observed on the TECS during tilting and holding. Figure 2 shows cropped images of the mouse embryos in motion. Between each 10 degree tilt (M1 and M3 in Figure 1B), the TECS plate was tilted at 1 degree/s. The motion of the medium was observed in M1 and M3 (Figure 1B). The embryo moved at a velocity of 1.5 mm/min on average (Figure 2A and B), which gave rise to shear stress of 7.0 x 10-3 dyn/cm2 at the bottom of the centre in the microdrop at a velocity of 0.1 mm/min. When the TECS plate was held at a 10 degree tilt (M2 and M4 in Figure 1B), the embryo slid at a velocity of 0.03 mm/min (Figure 2C, D), which gave rise to a shear stress of 1.5 · 10–4 dyn/cm2 in the medium, where the motion of medium at the centre of the bottom was neglected in the motion phases. These values are summarized in Table 2.
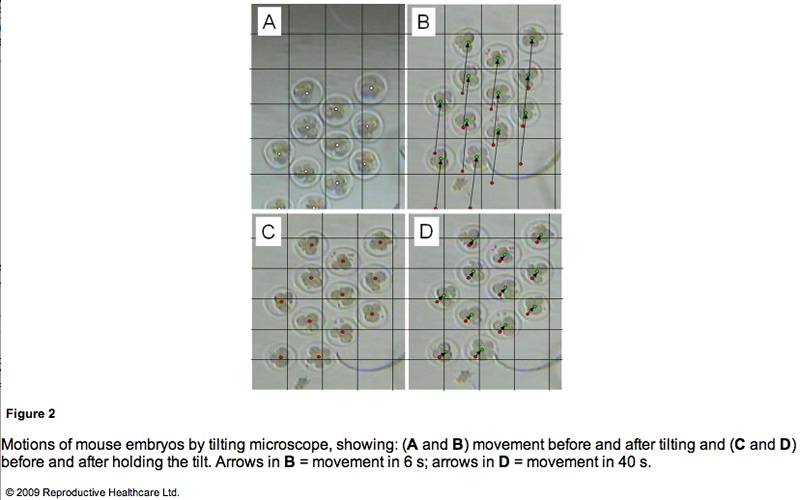
Mouse embryo development
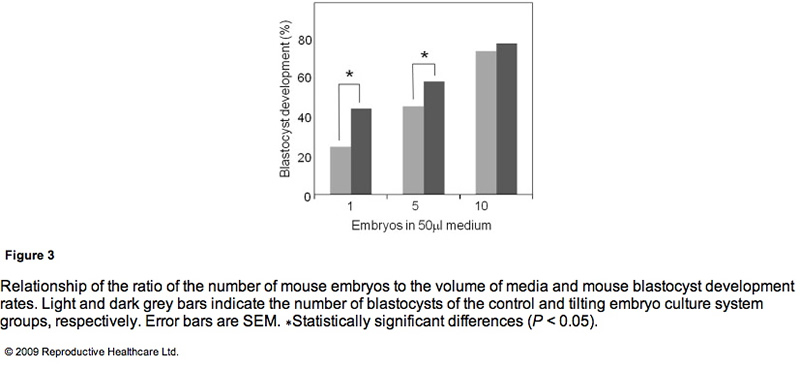
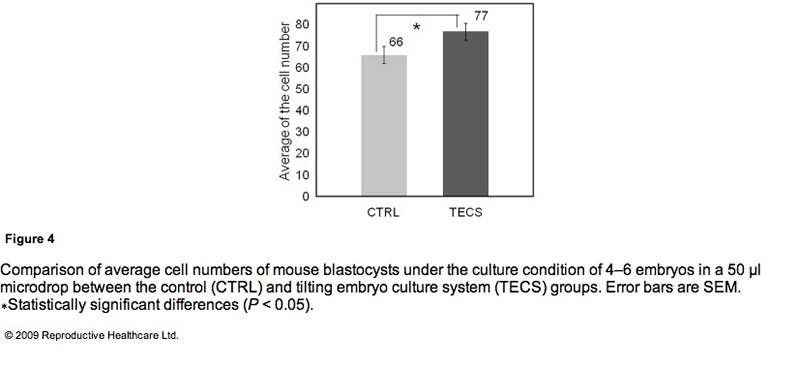
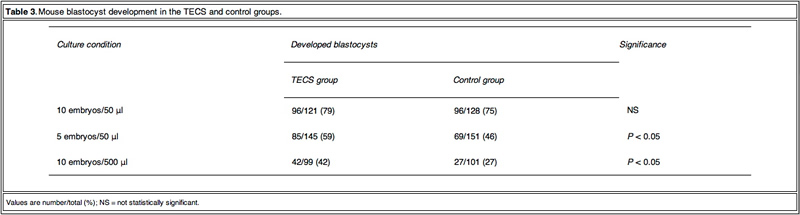
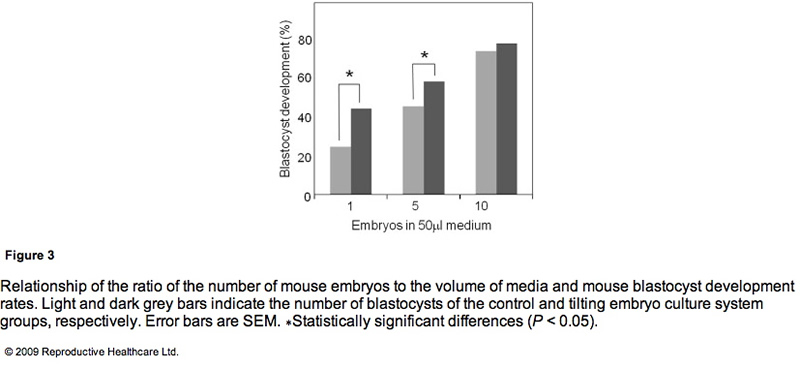
Table 3 shows the blastocyst development rates from 2- cell-stage mouse embryos cultured under several conditions. As shown in Figure 3, the blastocyst development rate was 79% for 10 embryos in a 50 ll microdrop using the TECS, which was not significantly different to the static culture controls (75%). Because mouse blastocyst development rate is reported to be influenced by the number of embryos in the culture medium (Melin et al., 2009) and in order to enhance the statistical difference between TECS and control, the number of embryos in the microdrop were deduced and the medium volume-to-embryo ratio, which might simulate poor development condition and lead to low-quality human embryos, was increased. The blastocyst development rate for four–six embryos in a 50 ll microdrop was less than that of 10 embryos, although in this poor condition, TECS significantly improved the blastocyst development rate (TECS 59% (n = 145) versus control 46% (n = 151); P < 0.05). When 10 embryos were cultured in 500 ll medium, the blastocyst development rate decreased to 27% (n = 101) in the static control, whereas for those cultured in the TECS it was significantly higher (42% (n = 99); P < 0.05)). The number of cells in the blastocysts cultured using the TECS was (mean ± SEM) 77 ± 4 cells (n = 34), while that of the control was 66 ± 4 cells (n = 26), as shown in Figure 4. There was a significant difference in the average cell number between the two groups (P < 0.05).

Human embryo development
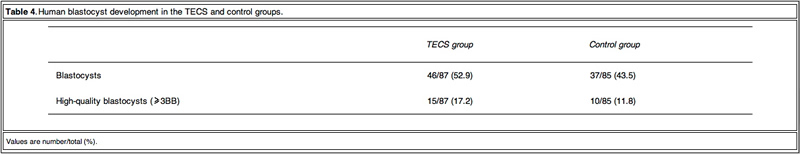
As TECS enhanced blastocyst development rate in mouse embryos, experiments were then conducted on thawed human embryos. The development rates to the blastocyst stage of the TECS and control groups were 53% and 45%, respectively (Table 4).
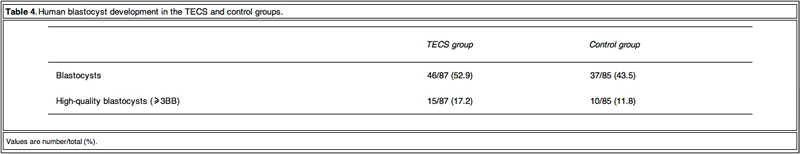
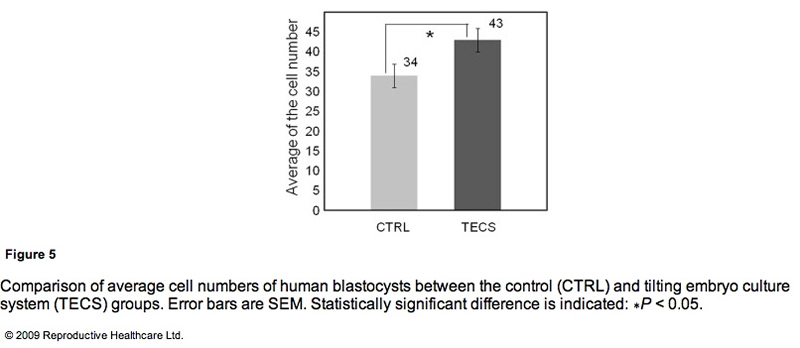
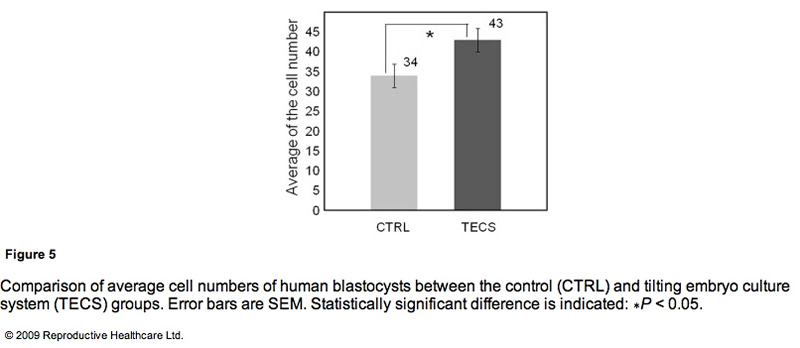
Due to the insufficient sample number of thawed human embryos, the study investigated the cell number of the developed blastocysts instead of the blastocyst development rate. Figure 5 shows the comparison of average cell numbers in human blastocysts developed by the TECS and control groups. The mean cell number of developed blastocysts at day 5 by the TECS was 43±3 cells (n=24), while that of the control was 34 ± 3 cells (n = 18). There was a significant difference in the average cell number between the two groups (P < 0.05).
Discussion
The Fallopian tube is a multifunctional organ, involved in receiving ovulated oocytes, providing a suitable environment for fertilization and early development and transporting embryos to the uterus. Here, light is shed on the mechanical properties of the Fallopian tube that might influence early development of embryos during culture. Early studies showed that the Fallopian tube is a mechanically active organ and may have influence on development due to: (i) shear stress by a tubal fluid flow; (ii) compression by peristaltic tubal wall movement; (iii) buoyancy; and (iv) kinetic friction force between embryo and cilia. Punctuated velocities of maxima from 0.39 to 1.8 mm/min have been observed in rat oviducts for microspheres emulating the size of embryos (Xie et al., 2006). Inappropriate culture conditions could be detrimental. Indeed, embryos sense shear stress and development is compromised (Xie et al., 2006, Xie et al., 2007). Because shear stress is a function of the velocity of the embryo and the flow and because the contribution is important, it is propose that the velocity of embryos should be made similar to those in the oviduct. Similarity in the velocities of the mouse embryos was found but those of other species were not observed. The velocities in-vivo and ex-vivo should be investigated for optimization.
The observed velocity of mouse embryos on a TECS culture plate was of a similar order to those proposed in the oviduct. The embryo motion in the microdrops on the TECS plate relates directly to the plate motion. The observed embryo images on the TECS plate suggest that the velocities and shear stresses of embryos (Table 2) in the TECS plate and the oviduct are similar. This comparative experiment suggests that the TECS can apply physiological mechanical stimuli to mouse embryos. In the case of human embryos, the embryo velocity in the microdrop on the TECS plate was the same order (approximately 1 mm/min) as those of mouse embryos and below 1.2 dyn/cm2. Therefore, the TECS could apply physiological stimuli without an excess amount of shear stress that might cause damage to the embryos. Furthermore, the detrimental or beneficial shear stress would be different at each developmental stage and future experiments will address this.
Embryo motion in fluid is affected not only by species differences but also the environment of the embryos. For example, when cumulus cells are coated with zona, embryos with the cells stick on the bottom of the dish. To optimize the parameters (θr and Vr) for each embryo condition, the relationship between the motions in fluid and physical characteristics of embryos (such as density, dimension and zona surface structure) should be considered.
The TECS significantly improved development in low-quality human embryos and suboptimal culture conditions in mouse embryos. The significance of the improvement by the TECS was dependent on the number of mouse embryos in the medium. As mentioned in the results, the blastocyst development rate significantly improved in the case of four–six mouse embryos in the microdrop. In the case of 10 embryos in the microdrop, blastocyst development rates of the TECS and control groups were 75% and 79%, respectively, and the difference was not significant. A higher number of embryos in the medium can improve the blastocyst development rate. According to previous reports that the concentration and production of autocrine and/or paracrine factors enhance mouse embryo development (Contramaestre et al., 2008, Kawamura et al., 2005), a higher number of embryos in the medium can improve the blastocyst development rate. The results in Figure 3 demonstrate that blastocyst development rate was significantly improved by the TECS when using a lower ratio of mouse embryo to media volume. This may be due to diffusion of growth factors and/or waste products, which would be facilitated by the TECS motion in the mouse embryo culture. However, mechanical stimuli could also be beneficial. Indeed, in bone and endothelial cells, down-stream transcription factors in the nucleus have been shown to be activated by mechanical stimuli, such as shear stress and mechanotransduction, and gene transcription and DNA syntheses were also activated (Wang and Thampatty, 2006). Due to the enhancement of cell division induced by these activations, cell numbers would be increased without apoptosis.
The results demonstrated that the cell numbers of the mouse and human blastocysts cultured in the TECS were greater than those cultured under control conditions and that TECS can improve the quality of those blastocysts. The increase in cell number of the mouse and human blastocysts suggests that TECS could enhance cell division of human embryos. According to published material (Cui et al., 2008, Xie et al., 2006), negative correlations between the percentage of TUNEL-positive cells and cell numbers in mouse embryos have been suggested. Therefore, although not measured, reduction of necrosis and/or apoptosis may be one explanation for the increased cell number in blastocysts obtained from TECS culture. Future experiments will confirm or refute this theory.
Finally, there are advantages of the TECS in clinical use. Although the culture conditions are different in each assisted reproduction laboratory, a benefit of this system is its ability to be rapidly implemented because it can be adapted to multiple styles of culture dishes/approaches. These results of thawed mouse and human embryo development indicate that the clinical study of embryo culture using the TECS can be extended without problems. To demonstrate the clinical importance of the TECS in human embryo development, a clinical multicentre study is being prepared on human embryo development using the TECS and embryo transfer. The improved quality of developed embryos by the TECS might contribute to enhanced pregnancy rates in clinical practice. As blastocyst cell numbers were increased by the TECS, pregnancy rates resulting from embryos cultured in this system might be improved.
In conclusion, the TECS enhanced blastocyst development rates of mouse embryos after the 2-cell stage and caused a significant increase of cell number in blastocysts. Thawed human embryos after the 3-cell stage tend to show an improved blastocyst development rate when cultured by the TECS. In particular, the improvements made by the TECS were significant in low-quality embryos and suboptimal culture conditions. One possible reason for the improvements could be mechanical stimuli by embryo motion based on the comparison of both mouse and human embryo development results.
Acknowledgements
This study was supported by a grant-in-aid for Scientific Research on Priority Areas (No. 17076006 to K.N.) and Special
Coordination Funds for Promoting Sciences and Technology from the Ministry of Education, Science, Sports, and Culture, Japan (to K.M.).
References
Cabrera, L.M., Heo, Y.S., Ding, J., et al., 2006. Improved blastocyst development with microfluidics and Braille pin actuator enabled dynamic culture. Fertil. Steril., S-43.
Cohen, J., Simons, R.F., Edwards, R.G., et al., 1985. Pregnancies following the frozen storage of expanding human blastocysts. J In Vitro Fert. Embryo Transfer 2, 59–64.
Contramaestre, A.P., Sifontes, F., Marin, R., et al., 2008. Secretion of stem cell factor and granulocyte-macrophage colony-stimulating factor by mouse embryos in culture: influence of group culture. Zygote 16, 297–301.
Cui, X.S., Shen, X.H., Kim, N.H., 2008. High motility group box 1 (HMGB1) is implicated in preimplantation embryo development in the mouse. Reprod. Domest. Anim. 75, 1290–1299.
Eddy, C.A., Pauerstein, C.J., 1980. Anatomy and physiology of the fallopian tube. Clin. Obstet. Gynecol. 23, 1177–1193.
Fox, G.J., 2007. The Mouse in Biomedical Research, second ed. Academic Press, pp. 97–98.
Gardner, D.K., Lane, M., Stevens, J., et al., 2000. Blastocyst score affects implantation and pregnancy outcome. Fertil. Steril. 73, 1155–1158.
Halbert, S.A., Tam, P.Y., Blandau, R.J., 1976. Egg Transport in the rabbit oviduct: the roles of cilia and muscle. Science 191, 1052–1053. Hardy, K., Handyside, A.H., Winston, R.M.L., 1989. The human
blastocyst: cell number, death and allocation during late preimplantation development in vitro. Development 107, 597–604. Kawamura, K., Fukuda, J., Kumagai, J., et al., 2005. Gonadotropin-releasing hormone I analog acts as an antiapoptotic factor in
mouse blastocysts. Endocrinology 146, 4105–4116. Lyons, R.A., Saridogan, E., Djahanbakhch, O., 2006. The reproductive significance of human fallopian tube cilia. Hum. Reprod. 12, 363–372.
Melin, J., Lee, A., Foygel, K., et al., 2009. In vitro embryo culture in defined sub-microliter volumes. Dev. Dyn. 238, 950–955.
Mottla, G.L., Adelman, M.R., Hall, J.L., et al., 1995. Lineage tracing demonstrates that blastomeres of early cleavage-stage human pre-embryos contribute to both trophectoderm and inner cell mass. Hum. Reprod. 10, 384–391.
Nagai, T., Ding, J., Moor, R.M., 1993. Effect of follicle cells and steroidogenesis on maturation and fertilization in vitro of pig oocytes. J. Exp. Zool. 266, 146–151.
Staigmiller, R.B., Moor, R.M., 1984. Effect of follicle cell on the maturation and developmental competence of ovine oocytes matured outside the follicle. Gamete Res. 9, 221–229.
Suarez, S.S., 1987. Sperm transport and motility in the mouse oviduct: Observation in situ. Biol. Reprod. 36, 203–210.
Wang, J.H.C., Thampatty, B.P., 2006. An introductory review of cell mechanobiology. Biomech. Model. Mechanobiol. 5, 1–16.
Xie, Y., Wang, Y., Zhong, W., et al., 2006. Shear stress induces preimplantation embryo death that is delayed by the zona pellucida and associated with stress-activated protein kinase mediated apoptosis. Biol. Reprod. 75, 45–55.
Xie, Y., Wang, Y., Puscheck, E.E., et al., 2007. Pipetting causes shear stress and elevation of phosphorylated stress-activated protein kinase/jun kinase in preimplantation embryos. Mol. Reprod. Dev. 74, 1287–1294.
Zervomanolakis, I., Ott, H.W., Hadziomerovic, D., et al., 2007. Physiology of upward transport in the human female genital tract. N.Y. Acad. Sci. 1101, 1–20.
Declaration: Keiji Narusee launched the bio-venture company, Strex Inc., in 2003 and serves as a Chief Scientific Officer. The other authors report no financial or commercial conflicts of interest.
Received 16 May 2009; refereed 8 June 2009; accepted 11 November 2009.
http://www.rbmojournal.com/article/PIIS1472648309002727/fulltext

 (2)
(2)
9334




 Click on image for full size
Click on image for full size